Virale Eukaryogenese
Virale Eukaryogenese (englisch viral eukaryogenesis) ist die Bezeichnung einer Hypothese, nach der sich der Zellkern eukaryotischer Lebensformen aus einem großen DNA-Virus in im Zuge einer Endosymbiose in einem methanogenen Archaeon oder einem Bakterium entwickelt hat. Nach diesem Szenario entwickelte sich das Virus später zum eukaryotischen Zellkern, indem es Gene aus dem Wirtsgenom übernahm und schließlich dessen Rolle an sich riss. Die Hypothese wurde erstmals 2001 von Philip Bell vorgeschlagen.[1] Sie bekam weitere Unterstützung durch die Entdeckung großer, komplexer DNA-Viren (wie dem Mimivirus), die zur Proteinbiosynthese fähig sind.
Die virale Eukaryogenese wird aus mehreren Gründen kontrovers diskutiert. Zum einen wird gelegentlich argumentiert, dass die behaupteten Beweise für den viralen Ursprung des Zellkerns auch im Umkehrschluss für den nuklearen Ursprung einiger Viren verwendet werden können.[2] Zum anderen hat diese Hypothese die seit langem geführte Debatte darüber, ob Viren lebende Organismen sind, weiter angeheizt.[2]
Hypothese
Die Hypothese der viralen Eukaryogenese besagt, dass Eukaryoten aus drei ursprünglichen Komponenten bestehen: einem Virus-Anteil, der zum heutigen Zellkern wurde; einer prokaryotischen Zelle (nach der Eozyten-Hypothese ein Archaeon), die das Zytoplasma und die Zellmembran der modernen Zellen beisteuerte; und einer weiteren prokaryotischen Zelle (ein Bakterium), die durch Endozytose zum modernen Mitochondrium wurde.
Die Hypothese wurde mit dieser Bezeichnung erstmal von Philip John Livingstone Bell 2001 vorgeschlagen[1] Um 2005/2006 schlugen Forscher um Jean-Michel Claverie und Patrick Forterre darüber hinausgehend vor, dass der Übergang von RNA- zu DNA-Genomen zuerst in der Welt der Viren (Virosphäre) stattgefunden hat.[3][4] Im Einklang mit der RNA-Welt-Hypothese hätten zelluläre Organismen danach zunächst als sog. Ribozyten (oder Ribozellen) ihre Erbinformation in einem RNA-Genom gespeichert. Ein DNA-basiertes Virus könnte dann seinen Ribozyten-Wirt dazu befähigt haben, seine Erbinformation von da an in der Form eines DNA-Genoms zu speichern, so wie dies heute bei allen bekannten zellulären Organismen der Fall ist.[2] Möglicherweise haben Viren als erste begonnen, ihre Erbinformation in DNA zu speichern, um diese vor RNA-abbauenden Enzymen der Wirtszellen zu schützen. Damit wäre der Beitrag von DNA-Viren zur Evolution zellulärer Organismen ebenso bedeutend gewesen sein wie der angenommene Beitrag von Alphaproteobakterien (als Vorläufer der Mitochondrien) oder von Cyanobakterien (als Vorläufer der Chloroplasten). Der ursprünglichen Hypothese folgend hätten die ersten Vertreter der drei Domänen – Archaeen, Bakterien und Eukaryoten die Fähigkeit zur Speicherung ihrer Erbinformation in einem DNA-Genom jeweils separat von einem anderen Virus erhalten.[4]
Über Ursprünge der DNA oder die Ursprünge der Reproduktion bei in Pro- und Eukaryotenzellen ist wenig bekannt. Es ist daher möglich, dass Viren an der Entstehung der ersten Zellen auf der Erde beteiligt waren.[5] Nick Lane begründete 2015, warum sich Bakterien und Archaeen mit Hilfe von Viren (möglicherweise Retroviren) unabhängig voneinander aus Ribozellen entwickelt haben könnten – ihre DNA-Replikationssysteme sind sehr verschieden (das der Eukaryoten ist dem der Archaeen vergleichsweise ähnlicher).[6]
Ursprünglich war insbesondere auch von einer Ribozelle als Vorfahr der Eukaryoten ausgegangen, auch wenn diese komplexer aufgebaut und fähig zur RNA-Verarbeitung gedacht war. Das diese ursprüngliche Variante eine Entstehung der Eukaryoten unabhängig von den Archaeen postulierte, war sie noch im Einklang mit ursprünglichen Überlegungen von Carl Woese und George Fox (1977), die solche hypothetischen Vorläufer der Eukaryoten als Urkaryoten bezeichneten.[7]
Nach heutigem Stand gilt jedoch die Abstammung der Eukaryoten von den Archaeen als wahrscheinlicher, wie sie zuerst von der Eozyten-Hypothese postuliert wurde. Im Laufe der Zeit fanden sich immer wieder neue Vertreter der Archaeen, die eine fortschreitende Nähe zu den Eukaryoten zeigten – angefangen mit den Crenarchaeota (TACK-Gruppe) über die Loki-Archaeen bis hin zu den Hodarchaeales unter den Heimdall-Archaeen (alle Asgard-Archaeen). Viren könnten aber auch in diesem Szenario einen entscheidenden Beitrag zur Eukaryogenese und damit zur Entstehung aller drei Lebensbereiche beigetragen zu haben (englisch out of virus hypothesis). Beispielsweise wurde vorgeschlagen, dass die Telomerase und Telomere als Schlüsselaspekte der eukaryotischen Zellreplikation, viralen Ursprungs sind. Darüber hinaus könnte der virale Ursprung des modernen eukaryotischen Zellkerns auf einer mehrfachen Infektion von Archaeen-Zellen, die bereits Bakterien als Mitochondrien-Vorläufer aufgenommen hatten, mit lysogenen Viren beruhen.[8]
Diskussion
Bezüglich des konkreten Ablaufs einer viralen Eukaryogenese sind mehrere theoretische Ansätze in der Diskussion.
Masaharu Takemura schlug 2001 ein Modell der viralen Eukaryogenese vor, bei dem sich ein dem modernen Pockenvirus ähnliches lysogenes Virus (Phylum Nucleocytoviricota) durch Genübernahme von bestehenden Bakterien- und Archaeenarten zu einem Zellkern entwickelt hat.[9] Dieses Virus übernahm nach dieser Vorstellung dann die Organisation des Erbguts der Zelle und wurde so zu ihrem Informationsspeicher, während die Zelle trotz des Eindringens des viralen Genoms ihre Fähigkeiten zur Gentranslation und andere allgemeinen Funktion behielt. In ähnlicher Weise behielt die an dieser Eukaryogenese beteiligte Bakterienart ihre Fähigkeit, Energie in Form von ATP zu produzieren, während sie gleichzeitig einen Großteil ihrer genetischen Information in den neuen vom Virus gestalteten Zellkern übertrug (endosymbiotischer Gentransfer). Es wird spekuliert, dass sich der moderne Zellzyklus, in dem Mitose, Meiose und Sexualität in allen Eukaryoten stattfinden, aufgrund eines viralen Gleichgewichts entwickelt haben, da die Viren typischerweise einem Kompromiss zwischen der Infektion möglichst vieler Wirte und der Tötung eines einzelnen Wirts durch virale Vermehrung folgen. Nach der Hypothese könnten die viralen Replikationszyklen denen von Plasmiden und mit lysogenen Phagen infizierten Bakterien entsprechen.
Takemura schlug 2020 vor, dass eine proto-eukaryotische Zelle von einem Vorfahren der heutigen NCLDV-Gattung Medusavirus parasitiert wurde. Die von den Medusaviren in der infizierten Zelle aufgebaute Virusfabrik zeigt eine deutliche Ähnlichkeit mit einem stark vereinfachten eukaryotischen Zellkern (d. h. einem in einer Lipidmembran eingeschlossenen DNA-Chromosom). Theoretisch könnte ein großes DNA-Virus die Kontrolle über eine Bakterien- oder Archaeen-Zelle übernommen haben. Anstatt sich zu vermehren und die Wirtszelle zu zerstören, würde es in der Zelle verbleiben (und so das oben erwähnte Dilemma zwischen der Infektion möglichst vieler Wirte und der Tötung eines einzelnen Wirts durch virale Vermehrung überwinden). Da das Virus die Kontrolle über die molekulare Maschinerie der Wirtszelle hat, würde es praktisch zu einem funktionalen Zellkern werden. Durch die Prozesse der Mitose und Zytokinese würde das Virus die gesamte Zelle als Symbiont rekrutieren – als ein neuer Weg, um zu überdauern und sich zu replizieren.[10]
Auch wenn Archaeenviren den modernen eukaryotischen Zellkernen evolutionär wahrscheinlich am ähnlichsten scheinen, sind nicht nur die Einzelheiten der Hypothese in der Diskussion, sie ist auch in Gänze nicht unumstritten. Weitere Experimente sind daher erforderlich, um offene Fragen zu klären.[11][12] Die Erforschung der 2022 erstmals per Metagenomik gefundenen Viren der Asgard-Archaeen (Asgardviren) steht noch ganz am Anfang. Eine große Schwierigkeit besteht in der Kultivierung dieser Archaeen, bis Februar 2023 dies erst bei zwei Vertretern der Asgard-Archaeen gelungen (Prometheoarchaeum syntrophicum und Lokiarchaeum ossiferum).
Ein Schlüssel zur Erforschung der Eukaryogenese konnte im Entwurf eines Stammbaums Chromatins und der eukaryotischen, archaealen und viralen[13] Histone liegen.[14]
Unterstützende Belege
Die Hypothese der viralen Eukaryogenese verweist auf den Zellzyklus von Eukaryoten, insbesondere auf Sexualität und Meiose, als Belege.[11] Der eukaryotische Zellkern enthält lineare DNA mit speziellen Endsequenzen, wie die mancher DNA-Viren (im Gegensatz zum zirkulären bakteriellen Genomen); er verwendet mRNA-Capping und hat eine räumliche Trennung von Transkription und Translation. Eukaryotische Zellkerne sind auch zur zytoplasmatischen Replikation fähig. Einige große Viren verfügen über eine eigene DNA-abhängige RNA-Polymerase.[2] Die Übertragung von „infektiösen“ Kernen wurde bei vielen parasitären Rotalgen dokumentiert.[15]
Zu den jüngeren Belegen der Hypothese gehört die Entdeckung durch Chaikeeratisak et al. im Jahr 2017, dass der Riesenbakteriophage 201Φ2-1 (Spezies Pseudomonas-Virus 201phi21, wiss. Serwervirus 201phi21, früher in der Gattung Phikzvirus alias phiKZ-like viruses, vom Morphotyp Myoviren[16][17][18]) bei der Infektion einer Bakterienzelle eine kernähnliche Struktur (Virusfabrik, VF) um die Region der Genomreplikation herum aufbaut. Transkription und Translation sind räumlich entkoppelt, d. h. die erzeugte virale mRNA wird in das Zytoplasma transportiert wird, wo sie der Translation unterzogen wird.[19] Dasselbe Team fand auch heraus, dass dieser Phage für ein eukaryotisches Homolog des Tubulins (PhuZ) kodiert. Dieses hat die Aufgabe die Virusfabrik während der Genomreplikation in der Mitte der Zelle zu positionieren.[20] Die PhuZ-Spindel teilt mehrere spezifische Eigenschaften mit eukaryotischen Spindeln: dynamische Instabilität, bipolare Filamentanordnungen und zentral positionierte DNA.[12] Darüber hinaus verfügen viele Klassen des Phylums Nucleocytoviricota (engl. nucleocytoplasmic large DNA viruses, NCLDVs) wie die Gattung Mimivirus über den Apparat zur Produktion von m7G-verkappter mRNA und enthalten Homologe des eukaryotischen Cap-Bindungs-Proteins eIF4E (engl. Eukaryotic translation initiation factor 4E). Zu Gunsten der viralen Eukaryogenese wird in diesem Zusammenhang auf das Fehlen dieser Merkmale bei Archaeen verwiesen. Es scheint immer noch zwischen den Archaeengruppen, die am nächsten mit den Eukaryoten verwandt sind, und den Eukaryoten selbst in Bezug auf den Zellkern eine beträchtliche Lücke zu klaffen. Angesichts dieser und anderer neuerer Erkenntnisse modifizierte Bell seine ursprüngliche These dahingehend, dass als ein möglicher Virus-Vorfahr des Zellkerns eher ein NCLDV-ähnliches Archaeenvirus als ein Pockenvirus in Frage kommt. Ein weiterer Beleg für diese Annahme ist, dass der an der Entkopplung von Transkription und Translation beteiligte m7G-Capping-Apparat sowohl in Eukaryoten als auch in den Mimiviridae vorhanden ist, nicht aber in Lokiarchaeota, die als die nahe Verwandten der Eukaryoten gelten, was auch durch eine phylogenetische Analyse des m7G-Capping-Wegs unterstützt wird.[12]
Asgardviren
Im Zusammenhang mit der Hypothese sind die Viren der Asgard-Archaeen (Asgardviren) von besonderem Interesse! da ihre Archaeenwirte den Eukaryoten am nächsten stehen. Die bisher vorgeschlagenen bzw. vom International Committee on Taxonomy of Viruses bereits offiziell bestätigten Asgardviren teilen sich grob in folgende Gruppen:[21]
- Viren mit Kopf-Schwanz-Aufbau: Klasse Caudoviricetes (bisherige sind vom Morphotyp Siphoviren) – diese befallen Bakterien und Archaeen
- Verdandiviridae
- Viren des Phylums Preplasmiviricota (bisherige gehören zu den Tectiliviricetes) – zu dieser Gruppe gehören die Polinton-artigen Viren (PLVs) und die Virophagen, von denen es Vertreter wie das Mavirus gibt, die sich ins Genom der Wirtszelle integrieren können.
- Skuldviridae
- spindel-, flaschen- und ellipsoidförmige Viren: Familien Bicaudaviridae, Fuselloviridae und Ovaliviridae – diese Gruppen umfassen ausschließlich Archaeenviren
- „Muninnvirus“, „Huginnvirus“
- „Wyrdviren“
- evtl. auch helikal-filamentöse Viren: Ordnung Ligamenvirales
Siehe auch
- Endogenes Retrovirus und endogene virale Elemente (EVEs)
- Paläovirologie
Weiterführende Literatur
- J. Durzyńska, A. Goździcka-Józefiak: Viruses and cells intertwined since the dawn of evolution. In: Virology Journal. 12. Jahrgang, Oktober 2015, S. 169, doi:10.1186/s12985-015-0400-7, PMID 26475454, PMC 4609113 (freier Volltext) – (englisch).
- H. L. Hendrickson, A. M. Poole: Manifold Routes to a Nucleus. In: Frontiers in Microbiology. 9. Jahrgang, 2018, S. 2604, doi:10.3389/fmicb.2018.02604, PMID 30416499, PMC 6212462 (freier Volltext) – (englisch).
- Patrick Forterre, M. Gaïa: Giant viruses and the origin of modern eukaryotes. In: Current Opinion in Microbiology. 31. Jahrgang, Juni 2016, S. 44–49, doi:10.1016/j.mib.2016.02.001, PMID 26894379 (englisch).
- Patrick Forterre: The origin of viruses and their possible roles in major evolutionary transitions. In: Virus Research. 117. Jahrgang, Nr. 1, April 2006, S. 5–16, doi:10.1016/j.virusres.2006.01.010, PMID 16476498 (englisch).
Einzelnachweise
- ↑ a b Philip John Livingstone Bell: Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus? In: Journal of Molecular Evolution. 53. Jahrgang, Nr. 3, September 2001, S. 251–6, doi:10.1007/s002390010215, PMID 11523012, bibcode:2001JMolE..53..251L (englisch).
- ↑ a b c d Jean-Michel Claverie: Viruses take center stage in cellular evolution. In: Genome Biology. 7. Jahrgang, Nr. 6, 16. Juni 2006, S. 110, doi:10.1186/gb-2006-7-6-110, PMID 16787527, PMC 1779534 (freier Volltext) – (englisch).
- ↑ Patrick Forterre: The two ages of the RNA world, and the transition to the DNA world: a story of viruses and cells. In: Biochimie, Band 87, Nr. 9–10, September–Oktober 2005, S. 793–803; doi:10.1016/j.biochi.2005.03.015, PMID 16164990.
- ↑ a b Patrick Forterre: Three RNA cells for ribosomal lineages and three DNA viruses to replicate their genomes: a hypothesis for the origin of cellular domain. In: Proceedings of the National Academy of Sciences of the United States of America. 103. Jahrgang, Nr. 10, März 2006, S. 3669–74, doi:10.1073/pnas.0510333103, PMID 16505372, PMC 1450140 (freier Volltext), bibcode:2006PNAS..103.3669F, JSTOR:30048645 (englisch).
- ↑ Jack T. Trevors: Genetic material in the early evolution of bacteria. In: Microbiological Research. 158. Jahrgang, Nr. 1, 29. September 2002, S. 1–6, doi:10.1078/0944-5013-00171, PMID 12608574 (englisch).
- ↑ Nick Lane: Der Funke des Lebens: Energie und Evolution, Konrad Theiss Verlag, Stuttgart 2017, ISBN 978-3-8062-3484-8. Originaltitel: The Vital Question: Why Is Life The Way It Is?, Profile Books, 2015; PDF (Memento vom 10. September 2017 im Internet Archive). Übersetzt von Martina Wiese, Monika Niehaus, Jorunn Wissmann. Dazu:
- Björn Lohmann: Aus energetischen Zwängen entstanden. spektrum.de, 24. Juni 2017, abgerufen am 8. Dezember 2022.
- Christmas 2015: The 6 best books in science. 27. November 2015, abgerufen am 8. Dezember 2022 (englisch).
- ↑ Carl R. Woese, George E. Fox: Phylogenetic structure of the prokaryotic domain: the primary kingdoms. In: Proceedings of the National Academy of Sciences. Band 74, Nummer 11, November 1977, S. 5088–5090; doi:10.1073/pnas.74.11.5088, PMID 270744, PMC 432104 (freier Volltext).
- ↑ Guenther Witzany: The viral origins of telomeres and telomerases and their important role in eukaryogenesis and genome maintenance. In: Biosemiotics. 1. Jahrgang, Nr. 2, 23. Juli 2008, S. 191–206, doi:10.1007/s12304-008-9018-0 (englisch, mitdenker.at [PDF]).
- ↑ Masaharu Takemura: Poxviruses and the origin of the eukaryotic nucleus. In: Journal of Molecular Evolution. 52. Jahrgang, Nr. 5, Mai 2001, S. 419–25, doi:10.1007/s002390010171, PMID 11443345, bibcode:2001JMolE..52..419T (englisch).
- ↑ Masaharu Takemura: Medusavirus Ancestor in a Proto-Eukaryotic Cell: Updating the Hypothesis for the Viral Origin of the Nucleus. In: Front. Microbiol. Band 11, S. 571831, 3. September 2020; doi: 10.3389/fmicb.2020.571831.
- ↑ a b Philip J. L. Bell: Sex and the eukaryotic cell cycle is consistent with a viral ancestry for the eukaryotic nucleus. In: Journal of Theoretical Biology. 243. Jahrgang, Nr. 1, November 2006, S. 54–63, doi:10.1016/j.jtbi.2006.05.015, PMID 16846615, bibcode:2006JThBi.243...54B (englisch).
- ↑ a b c Philip J. L. Bell: Evidence supporting a viral origin of the eukaryotic nucleus. In: Virus Research. 289. Jahrgang, 1. November 2020, S. 198168, doi:10.1016/j.virusres.2020.198168, PMID 32961211 (englisch).
- ↑ Paul B. Talbert, Karim-Jean Armache, Steven Henikoff: Viral histones: pickpocket’s prize or primordial progenitor? In: BMC Epigenetics & Chromatin, Band 15, Nr. 21, 28. Mai 2022; doi:10.1186/s13072-022-00454-7, PMID 35624484, PMC 9145170 (freier Volltext).
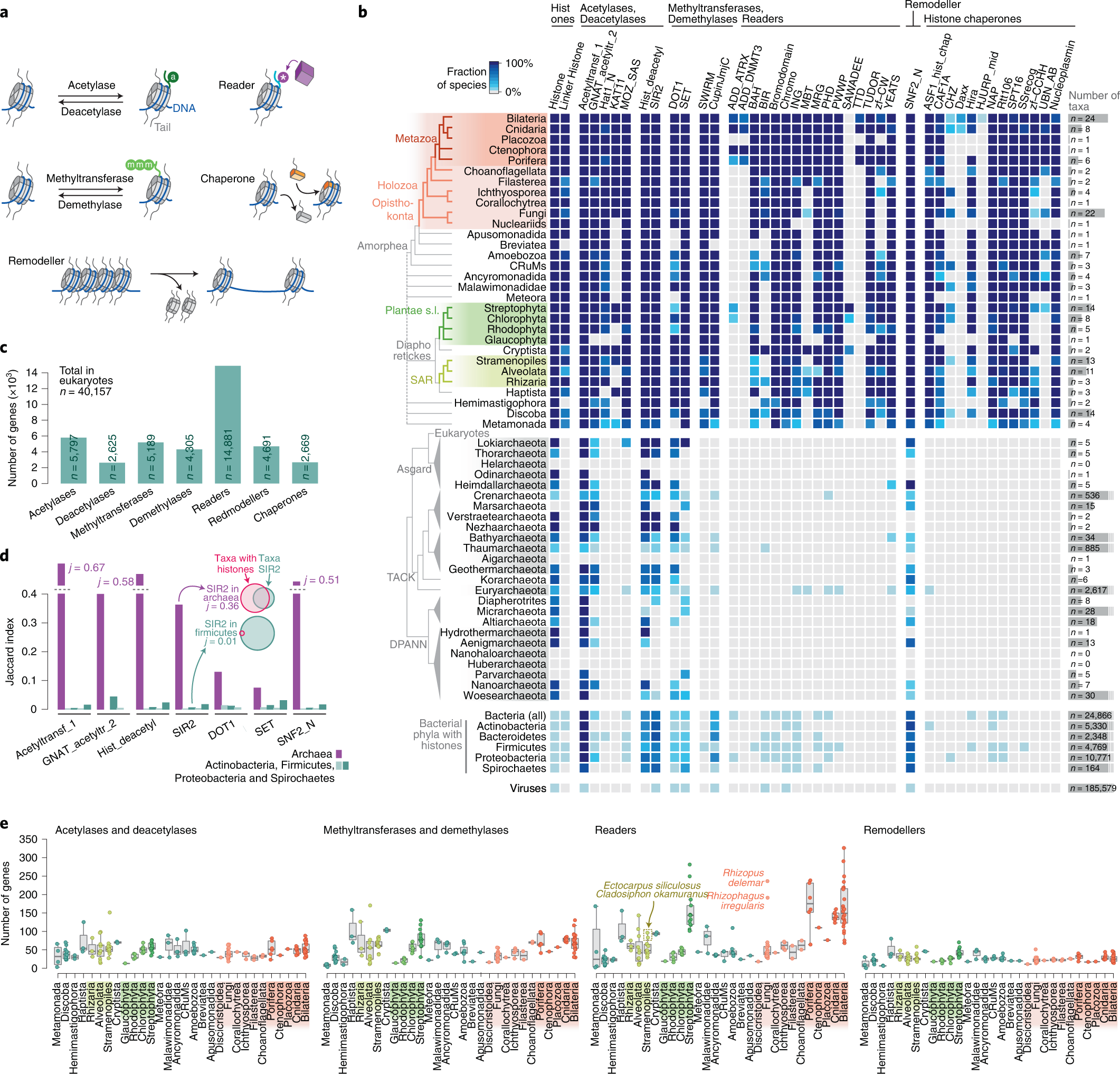
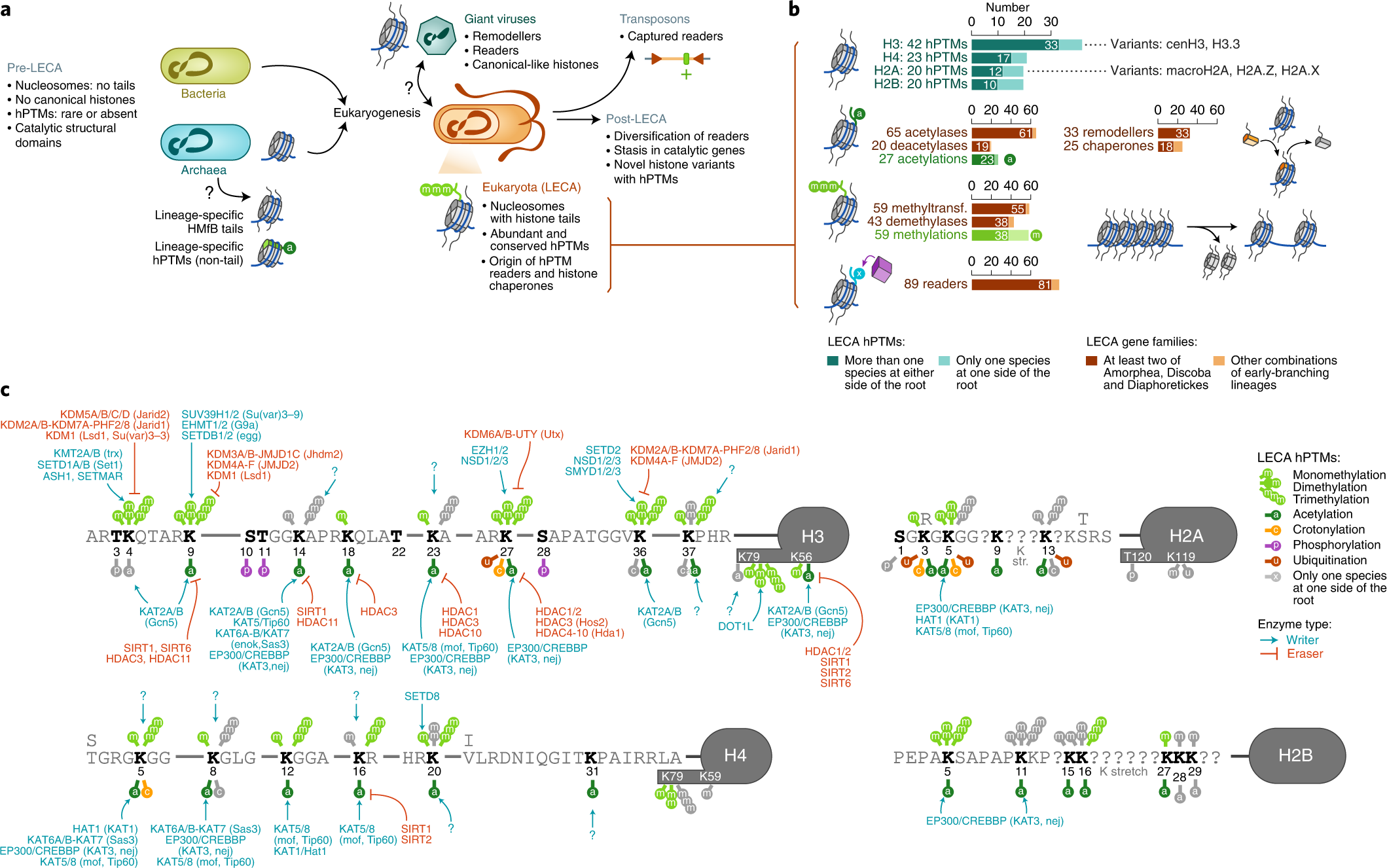
- ↑ Xavier Grau-Bové, Cristina Navarrete, Cristina Chiva, Thomas Pribasnig, Meritxell Antó, Guifré Torruella, Luis Javier Galindo, Bernd Franz Lang, David Moreira, Purificación López-Garcia, Iñaki Ruiz-Trillo, Christa Schleper, Eduard Sabidó, Arnau Sebé-Pedrós: A phylogenetic and proteomic reconstruction of eukaryotic chromatin evolution. In: Nature Ecology & Evolution, Band 6, S. 1007–1023, 9. Juni 2022; doi:10.1038/s41559-022-01771-6, PMID 35680998 PMC 7613034 (freier Volltext). Dazu:
- Fig. 1: Diversity of post-translational modifications in eukaryotic canonical and variant histones.
- Fig. 3: Taxonomic distribution of chromatin-associated gene classes.
- Fig. 6: Chromatin evolution and eukaryogenesis.
- Xavier Grau-Bové: Repository: Chromatin evolution. Auf: GitHub.
- Chromatin First Evolved in Ancient Microbes 1-2 Billion Years Ago, New Research Suggests. Auf: sci-news vom 13. Juni 2022.
- Chromatin originated in ancient microbes one to two billion years ago. Genomic and proteomic analysis reveals that the regulatory role of chromatin is a eukaryotic innovation. Auf: ScienceDaily vom 9. Juni 2022. Quelle: Center for Genomic Regulation (CRG)
- Chromatin originated in ancient microbes one to two billion years ago. Center for Genomic Regulation (CRG) vom 9. Juni 2022.
- ↑ Lynda J. Goff, Annette W. Coleman: Fate of Parasite and Host Organelle DNA during Cellular Transformation of Red Algae by Their Parasites. In: The Plant Cell. 7. Jahrgang, Nr. 11, November 1995, S. 1899–1911, doi:10.1105/tpc.7.11.1899, PMID 12242362, PMC 161048 (freier Volltext), JSTOR:3870197 (englisch).
- ↑ NCBI Taxonomy Browser: Pseudomonas phage 201phi2-1 (equivalent: Pseudomonas chlororaphis phage 201phi2-1).
- ↑ ICTV: Taxon Details: Genus: Phikzvirus, ursprünglicher Vorschlag: 2005.160-5B.v2.
- ↑ ICTV Vorschlag: 2022.066B Create five new genera of Pseudomonas jumbo phages (Caudoviricetes) (zip:docx).
- ↑ Vorrapon Chaikeeratisak, Katrina Nguyen, Kanika Khanna, Axel F. Brilot, Marcella L. Erb, Joanna K. C. Coker, Anastasia Vavilina, Gerald L. Newton, Robert Buschauer, Kit Pogliano, Elizabeth Villa, David A. Agard, Joe Pogliano: Assembly of a nucleus-like structure during viral replication in bacteria. In: Science. 355. Jahrgang, Nr. 6321, 13. Januar 2017, S. 194–197, doi:10.1126/science.aal2130, PMID 28082593, PMC 6028185 (freier Volltext), bibcode:2017Sci...355..194C (englisch).
- ↑ Vorrapon Chaikeeratisak, Katrina Nguyen, MacKennon E. Egan, Marcella L. Erb, Anastasia Vavilina, Joe Pogliano: The Phage Nucleus and Tubulin Spindle Are Conserved among Large Pseudomonas Phages. In: Cell Reports. 20. Jahrgang, Nr. 7, 15. August 2017, S. 1563–1571, doi:10.1016/j.celrep.2017.07.064, PMID 28813669, PMC 6028189 (freier Volltext) – (englisch).
- ↑ Tomas Alarcón-Schumacher, Susanne Erdmann: A trove of Asgard archaeal viruses. In: Nature Microbiology. Band 7, 27. Juni 2022, S. 931–932; doi:10.1038/s41564-022-01148-2. Dazu:
- Nicoletta Lanese: These Newly Discovered Viruses May Have Shaped The Rise of Complex Life on Earth. Auf: sciencealert vom 2. Juli 2022. Quelle: LiveScience.
{kind=link}
{kind=link}
{kind=link}