Eunotosaurus
| Eunotosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Replik eines Fossils von Eunotosaurus africanus mit erkennbar verbreiterten Rippenbögen | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Perm (Capitanium) | ||||||||||||
| 265,1 bis 259,9 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Eunotosaurus | ||||||||||||
| Seeley, 1892 | ||||||||||||
| Art | ||||||||||||
| ||||||||||||
Eunotosaurus ist eine Gattung reptilienartiger Sauropsiden aus dem mittleren Perm (265,1–259,9 mya) der Karoo-Supergruppe Südafrikas. Ihre Vertreter zeichneten sich durch verbreiterte und zahlenmäßig reduzierte Rippenbögen und einen rundlichen Körper aus. Die Gattung nimmt historisch wie aktuell eine bedeutende Rolle im Versuch des Verständnisses der Evolution der Schildkröten aus ihren prähistorischen Vorfahren ein.[1] Viele Fossilien wiesen einen halbstarren, schildkrötenähnlichen Brustkorb auf, der vermutlich eine Fortbewegung nach Art der Landschildkröten notwendig machte.[2]
Die Rippen waren breit und flach und berührten sich, so dass sie breite Platten vergleichbar dem Rückenschild einer Schildkröte bildeten. Außerdem waren die Anzahl, Größe und Aufbau der Wirbel weitgehend identisch mit denjenigen vieler Schildkrötenarten. Zudem schien Eunotosaurus einen fensterlosen (anapsiden) Schädel zu besitzen. Wegen dieser Eigenschaften wurde die Gattung oft zu den Anapsida bzw. Parareptilien gestellt und innerhalb dieser Gruppe als direkter Vorfahr der traditionell ebenfalls als Anapsiden betrachteten Schildkröten angesehen. Mit der spätestens aus molekulargenetischen Verwandtschaftsanalysen erwachsenen Erkenntnis, dass Schildkröten zu den Diapsiden gehören und einen sekundär anapsiden Schädel aufweisen, erschienen der Schädelbau, die verbreiterten Rippen und die ähnliche Wirbelsäule bei Eunotosaurus und Schildkröten jedoch nurmehr als ein Fall von konvergenter Evolution.[3] Neuere Untersuchungen erbrachten jedoch, dass Eunotosaurus einen reduzierten diapsiden Schädel besaß und seine Gemeinsamkeiten mit den Schildkröten auf Homologie statt auf Konvergenz beruhen.[4][5]

Fossilmaterial
Mehr als ein Jahrhundert nach der Erstbeschreibung war Eunotosaurus mit nicht einmal einem Dutzend Exemplaren und mit nur wenig überliefertem Material vom Schädel bekannt. Trotz der lückenhaften Überlieferung war es ausführlich beschrieben. Zwei zusätzliche Exemplare wurden 1999 in der Karoo-Supergruppe ausgegraben und beschrieben. Diese Fossilien sind nun im Bernard Price Institute for Palaeontological Research in Johannesburg und im Nationalmuseum von Bloemfontein ausgestellt. Obgleich relativ selten, ist Eunotosaurus in der Karoo-Supergruppe häufig genug, um als biostratigrafisches Marker herangezogen zu werden. Fossilien finden sich innerhalb der Beaufort-Gruppe in der oberen Tapinocephalus-Assemblage-Zone sowie der darüberliegenden Pristerognathus-Assemblage-Zone.[6]
Taxonomie und Systematik
Eunotosaurus wurde 1892 beschrieben, jedoch erst 1914 als Vorfahr der Chelonia, also der Ordnung der Schildkröten, vorgeschlagen. Der englische Zoologe D. M. S. Watson behauptete, dass Eunotosaurus eine Mosaikform zwischen Schildkröten und den Captorhinidae sei, die damals noch als Cotylosaurier bezeichnet wurden.[7] Er verglich Eunotosaurus mit "Archichelone", einem hypothetischen Vorfahren der Chelonia, wobei er anmerkte, dass seine Rippen intermediär sowohl zu denen der Schildkröten als auch zu denen anderer Tetrapoden waren. Watsons "Archichelone" besaß einen Beckengürtel, der auf der Wirbelsäule zurückgedrängt und unter dem Panzer platziert war. Allerdings zeigen die Fossilien von Eunotosaurus, dass sich sein Becken in einer für Tetrapoden normalen Stellung über den Rippen und nicht zwischen diesen, wie bei den modernen Schildkröten, befand.[1]
Bis in die späten vierziger Jahre galt Eunotosaurus gemeinhin als Vorfahr der Schildkröten. Der amerikanische Paläontologe Alfred Sherwood Romer behauptete 1956 in seinem Buch Osteology of the Reptiles aufgrund der vorhandenen Indizien, Eunotosaurus könne nicht zu den Chelonia gestellt werden. Stattdessen ordnete er Eunotosaurus den Anapsiden zu, innerhalb derer er jedoch eine unsichere Stellung einnahm.[1][8]
Eunotosaurus wurde 1954 seiner eigenen Familie, den Eunotosauridae, zugewiesen.[9] Diese Einteilung ist heute jedoch nicht mehr gebräuchlich. 1969 stellte man ihn in eine Unterordnung der Anapsiden, den Captorhinomorpha,[10] die heute innerhalb der Klade der Eureptilia angesiedelt wird.[11] Im Jahr 2000 wurde Eunotosaurus in die Klade der Parareptilien eingeordnet, wo er eine unabhängige Stellung von den Schildkröten und den Cotylosauriern einnimmt.[12] Eine phylogenetische Analyse der Parareptilien aus dem Jahr 2008 befand, dass Eunotosaurus ein Schwestertaxon der Milleretta sei und daher in die Familie der Millerettidae zu stellen sei.[13]
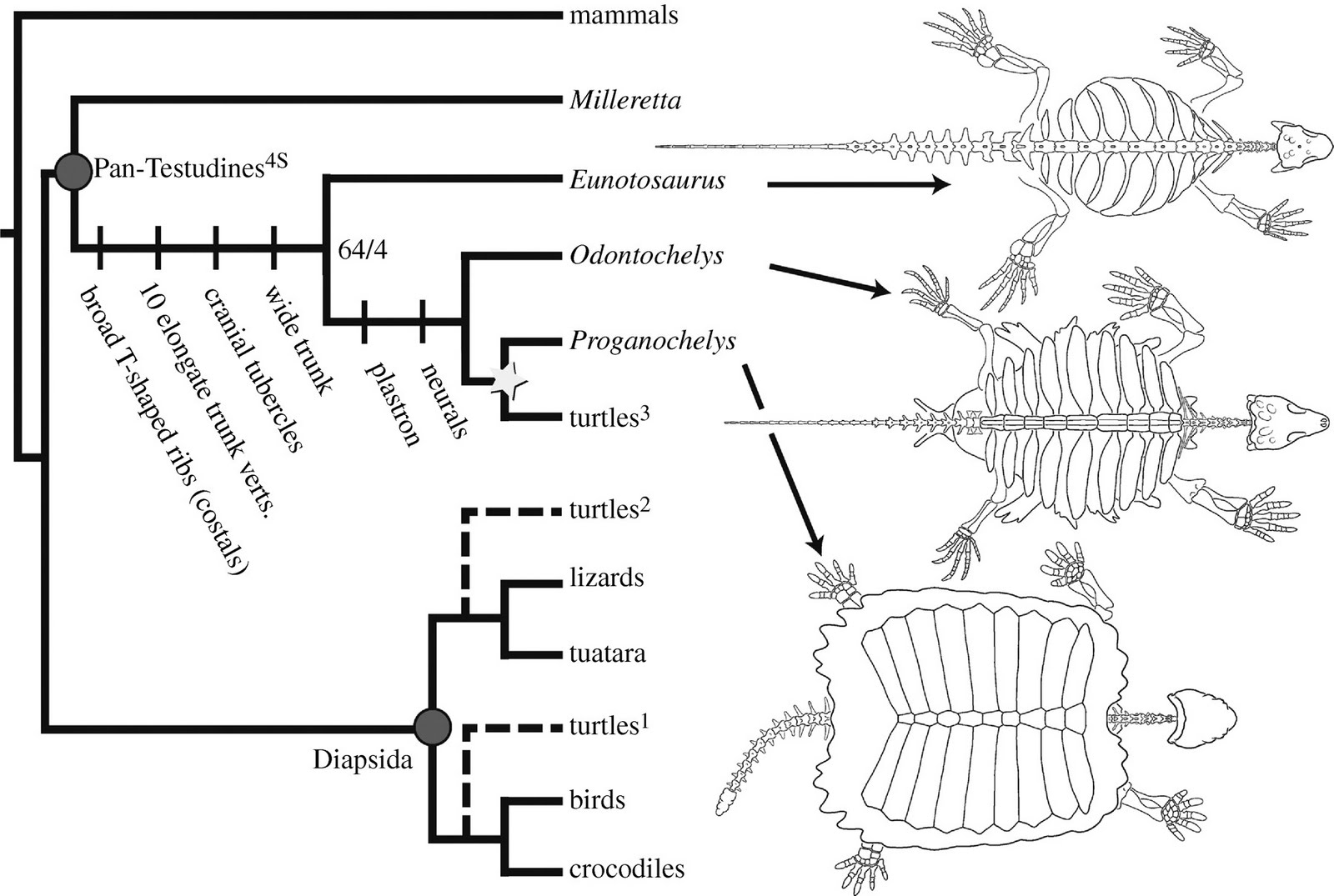
Eunotosaurus wurde in eine phylogenetische Analyse aus dem Jahr 2010 einbezogen, die der Frage nach dem Ursprung der Schildkröten nachging.[14] In neuerer Zeit werden Schildkröten auf der Grundlage genetischer und phylogenetischer Erkenntnisse als Diapsiden betrachtet und daher als näher verwandt mit den Eidechsen, Schlangen, Krokodilen und Vögeln als mit den Parareptilien oder allen anderen Anapsiden. Allerdings platziert der resultierende phylogenetische Baum die Schildkröten unter der Einbeziehung von Eunotosaurus und dem spättriassischen Proganochelys, einem Stammgruppenvertreter der Schildkröten, in einer Position, die der ursprünglichen Einordnung der Schildkröten als Anapsiden ähnelt. Die Studie behauptet, dass Eunotosaurus abgeleitete Merkmale der Rippen und der Rückenwirbel mit den frühesten Schildkröten teilte, was ihn zu einer Mosaikform mache. In der Studie wird eine Reihe von Merkmalen identifiziert, die es möglich machen, Eunotosaurus in einer echten Klade mit den Schildkröten zu vereinen. Diese gemeinsamen Merkmale beinhalteten breite T-förmige Rippen, zehn verlängerte Rumpfwirbel, kraniale Tuberkeln (kleine Höcker auf der Oberfläche des Schädels) und einen breiten Rumpf. Die Klade aus Eunotosaurus und den Schildkröten wurde als "Pan-Testudines" bezeichnet. Fortschrittlichere Vertreter der Pan-Testudines, wie z. B. Odontochelys, haben bereits einen Bauchpanzer.[14]
Unterstrichen wurde diese These durch eine Studie aus dem Jahr 2013, die die osteologische Feinstruktur der Knochen analysierte, mit der Ontogenese rezenter Schildkröten verglich und statistisch gewichtete. Sie kam zu dem Schluss, dass Eunotosaurus und die Schildkröten ein Monophylum bilden und innerhalb der Parareptilia stehen. Damit widersprachen beide Studien auch molekulargenetischen Untersuchungen, die die Schildkröten als Schwestergruppe der Archosaurier ausweisen.[15]
Eine weitere Studie aus dem Jahr 2014 zeigt, dass sich bei Eunotosaurus schon der eigenartige Atmungsmechanismus der Schildkröten entwickelte. Da die Rippen der Schildkröten in den Panzer integriert sind, sind sie unbeweglich, und für den Ein- und Ausstrom der Luft müssen spezielle Muskeln sorgen.[16] Im September 2015 veröffentlichte computertomographische Untersuchungen an verschiedenen Schädeln von Eunotosaurus ergaben, dass die Tiere zwei Schläfenfenster hinter den Augen besaßen, also einen diapsiden Schädel hatten. Dies ist besonders bei jungen Exemplaren deutlich zu sehen, bei älteren sind die Schläfenfenster dagegen fast vollständig verdeckt. Die Schädelontogenese von Eunotosaurus zeigt somit, wie sich aus einem diapsiden Schädel ein sekundär anapsider Schädel entwickelt, wie ihn die heutigen Schildkröten besitzen.[4] Möglicherweise verbreiterten sich die Rippen bei Eunotosaurus als Anpassung an eine grabende Lebensweise.[5]
Quellen
Literatur
- Tyler R. Lyson, Gabe S. Bever, Torsten M. Scheyer, Allison Y. Hsiang, Jacques A. Gauthier: Evolutionary Origin of the Turtle Shell. In: Current Biology. Bd. 23, Nr. 12, 2013, ISSN 0960-9822, S. 1113–1119, doi:10.1016/j.cub.2013.05.003.
Weblinks
Einzelnachweise
- ↑ a b c Ann Campbell Burke: The development and evolution of the turtle body plan: Inferring intrinsic aspects of the evolutionary process from experimental embryology. In: American Zoologist. Bd. 31, Nr. 4, 1991, ISSN 0003-1569, S. 616–627, doi:10.1093/icb/31.4.616.
- ↑ Stuart S. Sumida, Sean Modesto: A Phylogenetic Perspective on Locomotory Strategies in Early Amniotes. In: American Zoologist. Bd. 41, Nr. 3, 2001, S. 586–597, doi:10.1093/icb/41.3.586.
- ↑ Facts About Turtles: Eunotosaurus And Turtle Evolution. (Memento vom 12. September 2010 im Internet Archive) In: All-About-Reptiles.com. Abgerufen am 2. November 2014.
- ↑ a b G. S. Bever, Tyler R. Lyson, Daniel J. Field, Bhart-Anjan S. Bhullar: Evolutionary origin of the turtle skull. Nature. Bd. 525, 2015, S. 239–242, doi: 10.1038/nature14900
- ↑ a b Tyler R. Lyson, Bruce S. Rubidge, Torsten M. Scheyer, Kevin de Queiroz, Emma R. Schachner, Roger M.H. Smith, Jennifer Botha-Brink, G. S. Bever: Fossorial Origin of the Turtle Shell. Current Biology. Bd. 26, Nr. 14, 2016, S. 1887–1894, doi:10.1016/j.cub.2016.05.020
- ↑ Bruce S. Rubidge, Sean Modesto, Christian Sidor, Johann Welman: Eunotosaurus africanus from the Ecca–Beaufort contact in Northern Cape Province, South Africa — implications for Karoo Basin development. In: South African Journal of Science. Bd. 95, Nr. 11/12, 1999, ISSN 0038-2353, S. 553–555, Digitalisat (PDF; 1,11 MB) (Memento vom 26. Februar 2012 im Internet Archive).
- ↑ David M. S. Watson: Eunotosaurus africanus Seeley and the ancestors of the Chelonia. In: Proceedings of the Zoological Society of London. Bd. 84, Nr. 4, Article 53, 1914, ISSN 0370-2774, S. 1011–1020, doi:10.1111/j.1469-7998.1914.tb07724.x, Digitalisat.
- ↑ Alfred Sherwood Romer: Osteology of the Reptiles. University of Chicago Press, Chicago IL 1956, S. 772 (Reprint with new preface and taxonomic table. Krieger Publishing, Malabar FL 1997, ISBN 0-89464-985-X).
- ↑ Sidney H. Haughton, Adrian S. Brink: A bibliographical list of Reptilia from the Karoo Beds of Africa. In: Palaeontologia Africana. Bd. 2, 1954, ISSN 0078-8554, S. 1–187.
- ↑ Christopher Barry Cox: The problematic Permian reptile Eunotosaurus. In: Bulletin of the British Museum (Natural History). Geology. Bd. 18, Nr. 5, 1969, ISSN 0007-1471, S. 167–196, Digitalisat.
- ↑ Michel Laurin, Robert R. Reisz: A reevaluation of early amniote phylogeny. In: Zoological Journal of the Linnean Society. Bd. 113, Nr. 2, 1995, ISSN 0024-4082, S. 165–223, doi:10.1111/j.1096-3642.1995.tb00932.x.
- ↑ Sean P. Modesto: Eunotosaurus africanus and the Gondwanan ancestry of anapsid reptiles. In: Palaeontologia Africana. Bd. 36, 2000, S. 15–20.
- ↑ Juan Carlos Cisneros, Bruce S. Rubidge, Richard Mason: Analysis of millerettid parareptile relationships in the light of new material of Broomia perplexa Watson, 1914, from the Permian of South Africa. In: Journal of Systematic Palaeontology. Bd. 6, Nr. 4, 2008, ISSN 1477-2019, S. 453–462, doi:10.1017/S147720190800254X.
- ↑ a b Tyler R. Lyson, Gabe S. Bever, Bhart-Anjan S. Bhullar, Walter G. Joyce, Jacques A. Gauthier: Transitional fossils and the origin of turtles. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 6, Nr. 6, 2010, ISSN 0080-4649, S. 830–833, doi:10.1098/rsbl.2010.0371.
- ↑ Lyson 2013, S. 5–6.
- ↑ Tyler R. Lyson, Emma R. Schachner, Jennifer Botha-Brink, Torsten M. Scheyer, Markus Lambertz, G.S. Bever, Bruce Rubidge, Kevin de Queiroz (2014): Origin of the unique ventilatory apparatus of turtles, Nature Communications 5: 5211, DOI: 10.1038/ncomms6211
Auf dieser Seite verwendete Medien
Autor/Urheber: Smokeybjb, Lizenz: CC BY-SA 3.0
Life restoration of Eunotosaurus africanus.
- Based on skeletal in this cladogram, originally from "Transitional fossils and the origin of turtles" by Tyler R. Lyson, Gabe S. Bever, Bhart-Anjan S. Bhullar, Walter G. Joyce, and Jacques A. Gauthier (Proceedings of the Royal Society B).
{kind=link}